
Apresentaçőes
ORGANIZAÇĂO E SEGREGAÇĂO MEIÓTICA DOS CROMOSSOMOS SEXUAIS X1X2Y EM DUAS ESPÉCIES DE GRILOS DO GĘNERO ENEOPTERA (ORTHOPTERA, GRYLLIDAE) - PAULO ROBERTO BOEIRA FUCULO JUNIOR - - XXII CIC UFPel - 2013
ORGANIZAÇÃO E SEGREGAÇÃO MEIÓTICA DOS CROMOSSOMOS SEXUAIS X1X2Y EM DUAS ESPÉCIES DE GRILOS DO GÊNERO ENEOPTERA (ORTHOPTERA, GRYLLIDAE)
PAULO ROBERTO BOEIRA FUCULO JUNIOR¹; ANELISE FERNANDES E SILVA2; LUCIANO DE PINHO MARTINS²; EDISON ZEFA3
¹Universidade Federal de Pelotas – paulo.fuculo@hotmail.com
²Universidade Federal de Pelotas – anelise_fs@hotmail.com
²Instituto Nacional de Pesquisas da Amazônia - lucianodpm@gmail.com
³Universidade Federal de Pelotas – edzefa@gmail.com
INTRODUÇÃO
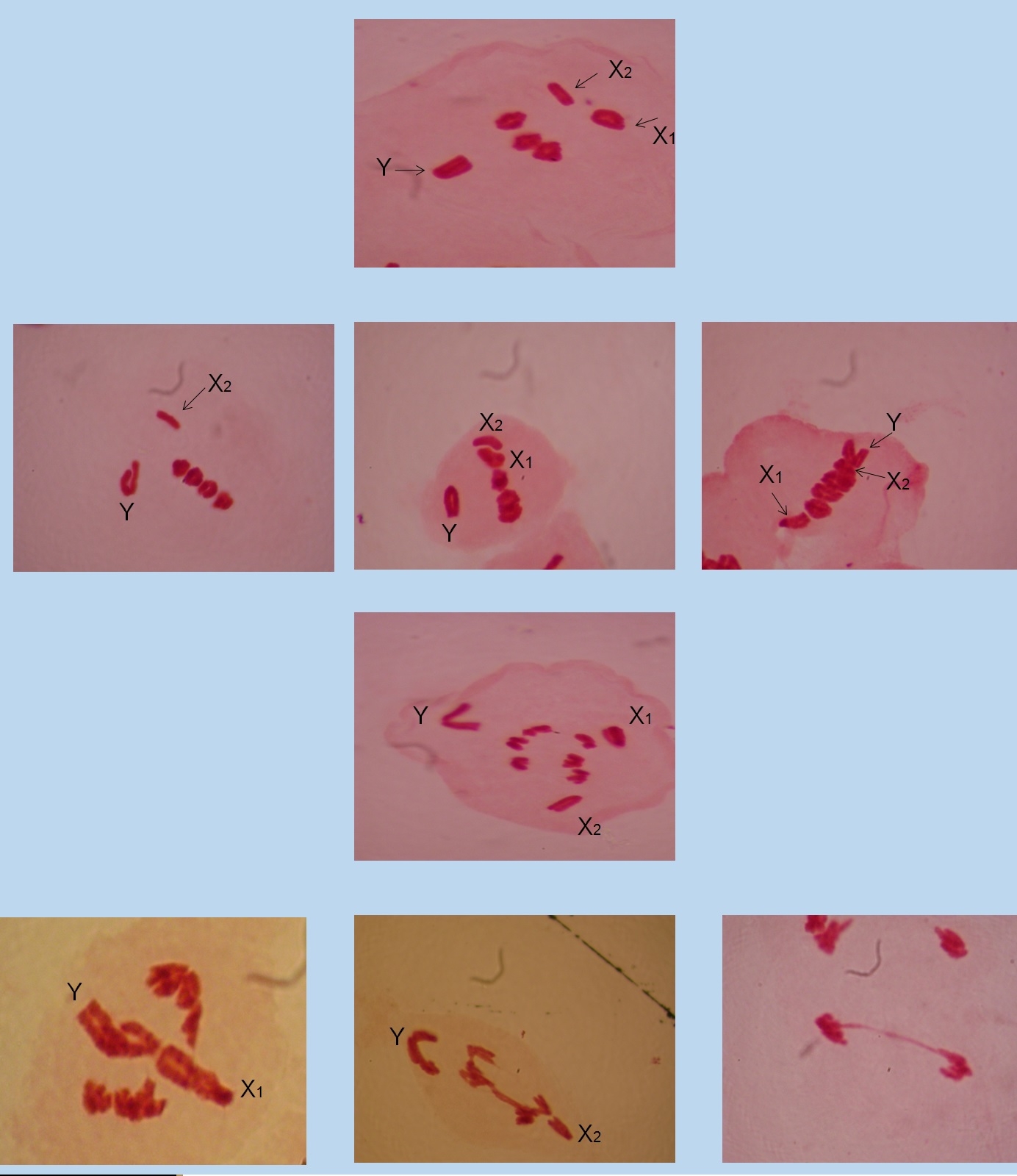
Os primeiros estudos cromossômicos em Eneoptera surinamensis (DE GEER, 1773) foram realizados por PIZA (1946), que determinou 2n = 9, XY1Y2 para os machos e 2n=8, XX para as fêmeas. Essa constituição cariotípica foi revista por CLAUS (1956) e MESA & BRAN (1964) que demonstraram que os machos apresentam 2n = 9, X1X2Y e as fêmeas 2n = 10, X1X1X2X2.
Os cromossomos autossomos de E. surinamensis formam bivalentes que se organizam na placa equatorial em função dos quiasmas, e segregam para pólos opostos durante a anáfase I. Os cromossomos sexuais X1X2Y, embora aquiasmáticos se orientam corretamente durante a metáfase I e anáfase I, migrando para os pólos opostos possibilitando a formação de 50% dos gametas com os cromossomos X1 e X2 e 50% com o cromossomo Y.
Dentre as espécies de Orthoptera e Mantodea que apresentam o mecanismo de determinação do sexo do tipo X1X2Y, o grau de pareamento dos cromossomos sexuais durante a meiose I é variável, e está relacionado ao tempo relativo de origem do sistema, de modo que quanto mais antigo for o rearranjo X/autossomos para a formação do mecanismo múltiplo de determinação do sexo, menor a frequência de quiasmas e o grau de pareamento desses cromossomos (WHITE, 1941; SAEZ, 1963; MESA et. al. 2001). Desta forma, em Orthoptera e Mantodea há espécies em que os cromossomos X1X2Y apresentam quiasmas e consequentemente formam trivalentes na Meiose I, e espécies em que o pareamento é reduzido ou totalmente ausente, como ocorre em E. surinamensis (WHITE, 1941; WHITE, 1973; MESA & BRAN, 1964).
Havendo ou não a formação de trivalentes na meiose I, os cromossomos X1 e X2 devem migrar para um dos pólos da célula e o cromossomo Y para o pólo oposto para que os gametas sejam viáveis, porém, nem sempre a segregação é correta. Em Scotussa daguerrei e Eurotettix lilloanus a formação de quiasmas e o pareamento dos cromossomos sexuais durante a meiose I é eventual e ocorrem problemas na orientação dos cromossomos durante a migração para pólos opostos, com conseqüente formação de gametas inviáveis (MESA, 1964; WHITE, 1941).
Em E. surinamensis não há nenhum tipo de pareamento ou quiasmas entre os cromossomos X1X2Y e também ocorrem problemas de migração incorreta com formação de gametas inviáveis (CLAUS, 1956; MESA & BRAN, 1964).
O objetivo deste trabalho foi caracterizar o comportamento dos cromossomos sexuais X1X2Y durante a meiose I em duas espécies de Eneoptera, destacando o processo de organização e segregação dos cromossomos e sexuais durante a metáfase I e anáfase I/telófase I.
METODOLOGIA
Os espécimes de E. surinamensis foram coletados em arbustos e serapilheira na Reserva Florestal Adolfo Ducke (Floresta Amazônica), município de Manaus, AM, em 08/11/2010 (03 indivíduos); no Parque Nacional de Monte Pascoal (Mata Atlântica), município de Itamaraju, BA em 15/07/2012 (01 indivíduo) e na Reserva Biológica de Duas Bocas (Mata Atlântica), município de Cariacica, ES em 21/07/2012 (02 indivíduos). Quatro indivíduos de Eneoptera sp. foram coletados no Parque Nacional de Monte Pascoal (Mata Atlântica), município de Itamaraju, BA em 15/07/2012.
A determinação taxonômica de E. surinamensis foi realizada de acordo com a morfologia dos escleritos fálicos (ROBILLARD & DESUTTER-GRANDCOLAS, 2005), além das características do som de chamado (MIYOSHI et al., 2007). A nova espécie de Eneoptera encontra-se em fase de descrição (Martins et al., em preparação).
As preparações citológicas foram obtidas a partir de testículos hipotonizados em solução de citrato de sódio (0.075M) por 5-10min, e fixados em Carnoy I (3 partes de álcool etílico e 1 parte de ácido acético glacial). O material foi macerado sobre lâmina com ácido acético 45% e as lâminas foram secadas ao ar livre, e coradas com orceína lacto-acético 0,5%.
Foram analisadas cinco lâminas de cada indivíduo e selecionadas as melhores fases da meiose I, sendo contabilizado o número total de metáfases I e anáfases I/telófases I em cada indivíduo, bem com o posicionamento e a distribuição dos cromossomos sexuais e autossomos.
RESULTADOS E DISCUSSÃO
Nos espécimes de E. surinamensis das três localidades foram observadas 94 metáfases I, 53 delas com os cromossomos sexuais X1 e X2 corretamente orientados para um pólo da célula e o cromossomo Y para o pólo oposto. Em 41 núcleos, os cromossomos sexuais X1 e/ou X2 e/ou Y apresentaram os centrômeros posicionados na placa equatorial, juntamente com os autossomos bivalentes.
Em E. surinamensis das três localidades ocorreram 73 anáfases I/telófases I. Em 35 delas, os cromossomos sexuais se orientaram normalmente, com o X1 e X2 para um dos pólos da célula e o cromossomo Y para o pólo oposto. Em 38 núcleos houve atraso na migração dos cromossomos X1 e/ou X2 e/ou Y, os quais se mantiveram na placa equatorial, enquanto os autossomos mostraram-se mais avançados rumo aos pólos, ou formaram pontes de cromatina entre os núcleos irmãos.
Nos espécimes de Eneoptera sp. foram registrados 111 núcleos em metáfase I, 70 deles com os cromossomos sexuais X1 e X2 corretamente orientados para um pólo da célula e o cromossomo Y para o pólo oposto. Em 41 núcleos, os cromossomos sexuais X1 e/ou X2 e/ou Y apresentaram os centrômeros posicionados na placa equatorial, juntamente com os autossomos bivalentes.
Nos mesmos espécimes de Eneoptera sp. foram observadas 26 anáfases I/telófase I. Em 20 delas os cromossomos sexuais se orientaram normalmente, com o X1 e X2 para um pólo da célula e o Y para o pólo oposto. Em seis núcleos houve atraso na migração dos cromossomos sexuais, com formação de pontes de cromatina, semelhante ao observado em E. Surinamensis.
Em E. surinamensis e Eneoptera sp., um número relativamente expressivo de núcleos apresentaram os cromossomos sexuais X1 e/ou X2 e/ou Y orientados na placa equatorial em função do centrômero durante a metáfase I. Esse comportamento é esperado para os cromossomos univalentes durante a metáfase II, em que os cromossomos se posicionam na placa equatorial em função dos centrômeros, permitindo que as cromátides irmãs migrem para pólos opostos durante a anáfase II.
O posicionamento dos cromossomos sexuais X1 e/ou X2 e/ou Y na placa equatorial durante a metáfase I sugere que seguirão para uma possível divisão equacional na anáfase I. Em casos em que há divisão equacional de cromossomos univalentes na anáfase I, os cinetocoros irmãos ficam dispostos numa orientação anfitélica para que ocorra divisão do centrômero e migração das cromátides irmãs para pólos opostos (MESA, 1984).
O processo de divisão equacional de cromossomos univalentes na anáfase I é mais lento quando comparado à separação dos cromossomos bivalentes, provavelmente devido aos processos enzimáticos envolvidos na separação dos centrômeros que unem as cromátides irmãs (SHAMU & MURRAY, 1992). Sugerimos que esse atraso seja o responsável pela formação das pontes de cromatina entre os núcleos telofásicos que ocorrem em E. surinamensis e Eneoptera sp.
O alinhamento dos cromossomos sexuais na placa equatorial e a migração tardia na Anáfase I, com formação de pontes de cromatina sugerem a formação de gametas inviáveis nessas espécies. Por outro lado, tanto E. surinamensis como Eneoptera sp. apresentam ampla distribuição geográfica (ZEFA, E. com. pess.), e compartilham a mesma estrutura cromossômica, preservando os mesmos fenômenos durante a meiose I e anáfase I/telófase I. Essas características sugerem o parentesco próximo entre as duas espécies e que esses fenômenos são relativamente conservados.
CONCLUSÕES
Os cromossomos sexuais X1X2Y de E. surinamensis são assinápticos e em alguns núcleos apresentam comportamento anômalo, incluindo o atraso na transição de metáfase I para anáfase I, segregação incorreta para os pólos e formação de pontes de cromatina entre núcleos anafásicos. Esse é o primeiro relato desse conjunto de anomalias meióticas presentes em duas espécies de Orthoptera proximamente relacionadas. Novos estudos serão necessários para compreender de modo esses anomalias influenciam no sucesso reprodutivo dessas espécies.
REFERÊNCIAS BIBLIOGRÁFICAS
Claus, G. 1956. La formule chromosomique du Gryllodea Eneoptera surinamensis De Geer et le comportement des chromosomes sexuels de cette espèce au cours de la spermatogenèse. Ann. Sc. Nat. Paris Zool. 11 (18): 63-106.
Mesa, A. 1984. The chromosomes of a relict species of eumastacid: Daguerreacris tandiliae Descamps & Liebermann, 1970 (Orthoptera, Eumastacoidea, Morseinae). Rev. Brasil. Genet. 7 (2): 219-229.
Mesa, A.; Bran, E.J. 1964. Acerca de los cromosomas de Eneoptera surinamensis. Anais do II Congresso Latino-Americano de Zoologia, São Paulo, 1: 9-16.
Mesa, A.; Fontanetti, C.S.; García-Novo, P. 2001. Does an x-autosome centric fusion in Acridoidea condemn the species to extinction? Journal of Orthoptera Research. 10 (2): 141-146.
Miyoshi, A.R.; Zefa, E., Martins, L.P., Dias, P.G.B.S. Drehmer, C.J., Dornelles, J.E.F. 2007. Stridulatory file and calling song of two populations of the tropical bush cricket Eneoptera surinamensis (Orthoptera, Gryllidae, Eneopterinae). Iheringia, Sér. Zool., 97 (4): 461-465.
Piza Jr, S.T. 1946. Uma nova modalidade de sexo-determinação no grilo Sul-Americano Eneopteras urinamensis. Na. Esc. Sup. Agric. “Luiz de Queiroz” Piracicaba, Brasil, 3: 69-88.
Robillard, T., Desutter-Grandcolas, L. 2005. A revision of Neotropical Eneopterinae crickets (Orthoptera, Grylloidea, Eneopteridae) with a phylogenetic discussion. Insect Systematics and Evolution 35 (4): 411-435.
Saez, F.A. 1963. Gradient of the heterochromatinization in the evolution of the sexual system Neo-X-Neo-Y.Portugaliae Acta Biologica, série A, 7 (1-2): 111-138.
Shamu, C. E.; Murray, A. W. 1992. Sister chromatid separation in frog egg extracts requires DNA topoisomerase II activity during anaphase. The Journal of Cell Biology, v. 117 (5), 921-934.
White, M.J.D. 1941. The evolution of the sex chromosome. I. The X0 and X1X2Y mechanisms in praying mantids. J. Genet. 42: 143-172.
White, M.J.D. Animal citology and evolution. 3.ed. London: Cambridge University Press, 1973. 961p.